
Primárna štruktúra bielkovín

Primárna štruktúra bielkovín je lineárna sekvencia aminokyselín v peptide alebo bielkovine.[1] Je to zoznam poradia aminokyselín, ktoré tvoria konkrétny polypeptid. Zapisuje sa vždy od N-konca po C-koniec, teda od aminoskupiny na jednej strane po karboxylovú skupinu na druhej strane. Biosyntéza bielkovín prebieha najčastejšie pomocou ribozómov v bunkách. Peptidy je možné pripraviť i v laboratóriu. Primárnu štruktúru bielkovín je možné priamo sekvenovať alebo určiť podľa príslušnej sekvencie DNA.
Vznik[upraviť | upraviť zdroj]
Biologický[upraviť | upraviť zdroj]
Aminokyseliny polymerizujú vďaka peptidovým väzbám, ktoré sú tvorené aminoskupinou jednej aminokyseliny a karboxylovou skupinou druhej aminokyseliny. Takto tvoria dlhý reťazec, z ktorého trčia do strán bočné reťazce aminokyselín. V biologických systémoch vznikajú bielkoviny v bunkách počas translácie na ribozómoch. Niektoré organizmy sú takisto schopné tvoriť krátke peptidy pomocou neribozomálnej syntézy peptidov, ktoré často využívajú iné aminokyseliny okrem štandardných 20 aminokyselín, a tieto peptidy môžu potom byť upravené, prepojené navzájom alebo uzavreté do cyklického reťazca.
Chemický[upraviť | upraviť zdroj]
Peptidy je možné syntetizovať chemicky pomocou mnohých laboratŕoynch metód. Chemické metódy typicky syntetizujú peptidu v opačnom poradí (od C-konca) oproti biologickej syntéze (ktorá prebieha od N-konca).
Zápis[upraviť | upraviť zdroj]
Sekvencia bielkovín je typicky zapisovaná ako zoznam písmen, z ktorých je bielkovina tvorená. Tento zápis začína N-koncom na ľavej strane a končí C-koncom na pravej strane. Buď sa používajú trojpísmenové skratky, alebo jednopísmenové skratky, ktoré predstavujú 20 bežných proteínogénnych aminokyselína niekedy sa používajú i špeciálne znaky, ktoré popisujú jednotlivé skupiny aminokyselín alebo nejednoznačné aminokyseliny (podobne, ako u zápisu nukleových kyselín).[1][2][3]
Peptidy je možné sekvenovať priamo alebo je možné ich sekvenovať z príslušnej sekvencie DNA. Existujú veľké databáze sekvencií, zbierajú známe sekvencie bielkovín.
| Aminokyselina | 3-písmenová skratka[4] |
1-písmenová skratka[4] |
|---|---|---|
| Alanín | Ala | A |
| Arginín | Arg | R |
| Asparagín | Asn | N |
| Aspartát | Asp | D |
| Cysteín | Cys | C |
| Glutamát | Glu | E |
| Glutamín | Gln | Q |
| Glycín | Gly | G |
| Histidín | His | H |
| Izoleucín | Ile | I |
| Leucín | Leu | L |
| Lyzín | Lys | K |
| Metionín | Met | M |
| Fenylalanín | Phe | F |
| Prolín | Pro | P |
| Serín | Ser | S |
| Treonín | Thr | T |
| Tryptofán | Trp | W |
| Tyrozín | Tyr | Y |
| Valín | Val | V |
Okrem týchto aminokyselín sa ešte niekedy vyskytujú dve aminokyseliny, a to pyrolyzín a selenocysteín:
- pyrolyzín sa označuje skratkami Pyl a O[5][6][7] a
- selenocysteín sa označuje skratkami Scy[5] alebo Sec[6][7] a U.
| Symbol | 3-písmenová
skratka |
Popis | Zahrnuté aminokyseliny |
|---|---|---|---|
| X | Xxx[5], Xaa[6], Unk[7] | Akákoľvek alebo neznáma aminokyselina | Všetky |
| B | Asx[6] | Aspartát alebo Asparagín | D, N |
| Z | Glx[6] | Glutamát alebo Glutamín | E, Q |
| J | Xle[6] | Leucín alebo Izoleucín | I, L |
| Φ | žiadna | Hydrofóbne | V, I, L, F, W, M |
| Ω | Aromatické | F, W, Y, H | |
| Ψ | Alifatické | V, I, L, M | |
| π | Malé | P, G, A, S | |
| ζ | Hydrofilné | S, T, H, N, Q, E, D, K, R, Y | |
| + | Kladne nabité | K, R, H | |
| - | Záporne nabité | D, E |
Úpravy[upraviť | upraviť zdroj]
Všeobecne sú polypeptidy lineárne nevetvené reťazce, takže ich primárna štruktúra sa dá popísať sekvenciou aminokyselín v ich reťazci. Niektoré bielkoviny však môžu byť spojené dokopy, najčastejšie pomocou disulfidových väzieb, takže ich primárna štruktúra potom vyžaduje upresnenie atómov, ktoré tvoria túto väzbu medzi jednotlivými bielkovinami, teda napríklad to, ktoré cysteíny tvoria disulfidovú väzbu. Medzi ďalšie podobné prípady patrí napríklad dezmozín.
Izomerizácia[upraviť | upraviť zdroj]
Chirálne centrá polypeptidového reťazca môžu podliehať racemizácii. Aj keď sa tým nemení sekvencia, menia sa tým chemické vlastnosti tejto sekvencie. U bielkovín môžu spontánne izomerizovať L-aminokyseliny na svojich Cα uhlíkoch, čím z nich vznikajú D-aminokyseliny, ktoré väčšina proteáz nevie štiepiť. Okrem toho môže prolín tvoriť stabilné trans-izoméry peptidovej väzby.
Posttranslačné úpravy[upraviť | upraviť zdroj]
Okrem uvedených úprav môžu byť niektoré aminokyseliny posttranslačne upravené. Tieto úpravy sú v krátkosti popísané nižšie.
N-koncová aminoskupina polypeptidu môže byť upravené kovalentne, môže tu prebehnúť napríklad

- acetylácia

- Kladný náboj na N-koncovej aminoskupine môže byť eliminovaný náhradou za acetylovú skupinu.
- formylácia

- N-koncový metionín sa často nachádza s formylovanou aminoskupinou, čiže má po translácii N-koniec blokovaný formylovou skupinou. Táto skupina (a niekedy i samotný metionín, ak ho nasleduje Gly alebo Ser) sa niekedy štiepi pomocou enzýmu deformylázy.
- tvorba pyroglutamátu
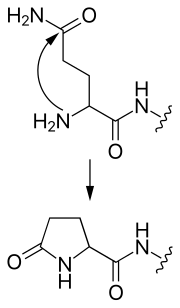
Tvorba pyroglutamátu z N-koncového glutamínu

- Glutamín na N-konci môže atakovať sám seba, čím vzniká cyklický pyroglutamátová skupina.
- myristoylácia

- Podobne ako u acetylácie môže byť viazaná myristoylová skupina, ktorá má namiesto metylu 14 hydrofóbnych vodíkov. Vďaka tomu je ideálna pre ukotvenie bielkovín do bunkovej membrány.
Podobne môže byť upravená C-koncová karboxylová skupina, kde môže prebehnúť napríklad
- aminácia

- Karboxylová skupina môže byť blokovaná amináciou, takže má neutrálny náboj namiesto záporného.
- naviazanie glykozylfosfatidylinozitolovej (GPI) kotvy
- Glykozylfosfatidylinozitol (GPI) je veľká, hydrofóbna fosfolipidová prostetická skupina, ktorá ukotvuje bielkoviny na bunkovú membránu. Pripája sa na polypeptidový C-koniec pomocou amidovej väzby, ktorá sa potom spája s etanolamínom, následne na sacharidy a nakoniec na fosfatidylinozitolový lipid.
Okrem týchto úprav môžu byť upravené i bočné reťazce aminokyselín. Tú môže prebehnúť napríklad
- fosforylácia
- Okrem štiepenia je fosforylácia asi tou najdôežitejšou chemickou modifikáciou bielkovín. Fosfátová skupina môže byť viazaná na hydroxylové skupiny serínu, treonínu a tyrozínu, čím pridáva záporný náboj na tomto mieste a tvorí neprirodzenú aminokyselinu. Fosforylácia bielkovín je katalyzovaná kinázami a opačná reakcia (defosforylácia) je katalyzovaná fosfatázami. Fosforylované tyrozíny sa často využívajú ako „úchytky“, zatiaľ čo fosforylovaný serín a treonín vyvolávajú konformačné zmeny, pravdepodobne kvôli novému zápornému náboju. Efekt fosforylácie serínu a treonínu sa niekedy sa simulovať mutáciou týchto aminokyselín na glutamát.
- glykozylácia
- Pod týmto názvom sa nachádza mnoho bežných a veľmi rozličných chemických úprav. Sacharidové skupiny môžu byť viazané na bočné reťazce serínu a treonínu alebo na bočnú amidovú skupinu asparagínu. Tieto úpravy majú mnoho funkcií, a to od zvýšenia rozpustnosti až po komplexné rozpoznávanie bielkovín. Všetky glykozylácie je možné blokovať pomocou niektorých inhibítorov, napríklad tunikamycínu.
- deamidácia (tvorba sukcinimidu)
- Pri tejto úprave napadá bočný reťazec asparagínu alebo aspartátu nasledujúcu peptidovú väzbu, čím vzniká symetrický sukcinimidový intermediát. Hydrolýza tohto intermediátu vedie k aspartátu alebo jeho β-aminokyselinovej verzii, izoaspartátu. (Izoaspartát má identický vzorec s aspartátom; jediný rozdiel je v tom, ktorá karboxylová skupina sa účastní hlavného polypeptidového reťazca.) V prípade asparagínu vedie tvorba oboch produktov k strate amidovej skupiny, preto sa táto úprava nazýva deamidácia.
- hydroxylácia
- Prolínové reziduá môžu byť hydroxylované na jednom z dvoch atómov. Podobne môže byť hydroxylovaný lyzín na jednom atóme. Hydroxyprolín je dôležitou súčasťou kolagénu, ktorý je bez neho nestabilný. Hydroxylácia je katalyzovaná enzýmom, ktorý vyžaduje kyselinu askorbovú (vitamín C). Nedostatky kyseliny askorbovej vedú k poruchám spojivového tkaniva, napríklad skorbutu.
- metylácia
- Niektoré reziduá môžu byť metylované, najčastejšie kladne nabité skupiny lyzínu a arginínu. Arginínové reziduá interagujú s fosfátovými skupinami nukleových kyselín a bežne tvoria vodíkové väzby s bázami, hlavne guanínom, v komplexoch proteín-DNA. Lyzínové reziduá môžu byť metylované až trikrát. Metylácia však nemení náboj na ich bočnom reťazci.
- acetylácia
- Acetylácia lyzínovej aminoskupiny je chemicky analogická k acetylácii N-koncovej aminoskupiny. Táto modifikácia sa však využíva na reguláciu viazania bielkovín na nukleové kyseliny. Vyrušenie kladného náboja lyzínu oslabuje elektrostatickú atrakciu k záporne nabitým nukleovým kyselinám.
- sulfácia
- Na hydroxylovú skupinu tyrozínu je možné naviazať sulfátovú skupinu. Táto modifikácia prebieha v Golgiho aparáte, zatiaľ čo ostatné prebiehajú v endoplazmatickom retikule. Podobne ako je to u fosforylovaných tyrozínov sa i sulfátované tyrozíny používajú na špecifické rozpoznávanie, napríklad v chemokinových receptoroch na povrchu bunky. Tak ako pri fosforylácii, naviazanie sulfátovej skupiny pridáva záporný náboj na neutrálny tyrozín.
- prenylácia a palmitoylácia

- Hydrofóbne izoprény (napríklad farnezylová, geranylový a geranylgeranylová skupina) a palmitoylová skupina môžu byť naviazané na sírový atóm cysteínových reziduí, čo umožňuje ich ukotvenie do bunkovej membrány. Na rozdiel od myristoylovej skupiny a GPI kotvy sa tieto skupiny nemusia pripájať na konce bielkoviny.
- karboxylácia
- Relatívne vzácna modifikácia, ktorá pridáva ďalšiu karboxylovú skupinu (a teda dvojitý záporný náboj) na bočný reťazec glutamátu, čím vzniká karboxyglutamát (Gla). Táto úprava slúži na posilnenie väzby na „tvrdé“ kovové ióny, napríklad vápnik.
- ADP-ribozylácia
- ADP-ribozylová skupina môže byť presunutá na niekoľko rôznych bočných reťazcov s rozličnými účinkami. Táto modifikácia je cieľom silných toxínov veľmi rozličných baktérií, napríklad Vibrio cholerae, Corynebacterium diphtheriae and Bordetella pertussis.
- ubikvitinylácia a SUMOylácia
- Rôzne dlhé poskladané proteíny môžu byť pripojené svojím C-koncom na aminoskupiny bočného reťazca lyzínu iných bielkovín. Ubikvitín je najbežnejším príkladom tejto úpravy a zvyčajne označuje bielkoviny určené na rozklad.
Väčšina úprav polypeptidového reťazca uvedená vyššie prebieha posttranslačne, teda až po tom, čo sa bielkovina syntetizuje na ribozóme. Zvyčajne prebiehajú v endoplazmatickom retikule, bunkovej organele eukaryotickej bunky.
Mnohé ďalšie chemické reakcie (napríklad naviazanie kyanoskupiny) môžu byť aplikované chemicky, ale nie sú prítomné v živých systémoch.
Štiepenie a spájanie[upraviť | upraviť zdroj]
Okrem úprav popísaných vyššie je najdôležitejšou úpravou primárnej štruktúry štiepenie peptidov (chemickou hydrolýzou alebo proteázami). Bielkoviny sa často syntetizujú v neaktívnej prekurzorovej forme - typicky časť na N-konci alebo C-konci blokuje aktívne miesto bielkoviny, čím bráni jej funkcii. Takýto proteín sa potom aktivuje štiepením inhibitujúceho peptidu.
Niektoré proteíny majú dokonca schopnosť toto štiepenie vykonať sami. Typicky je to vďaka hydroxylovej skupine serínu (vzácne treonínu) alebo tiolovej skupine cysteínu, ktorá napadá karbonylový uhlík predchádzajúcej peptidovej väzby, čím vzniká tetraédrický intermediát (hydroxyoxazidolidín (Ser/Thr) alebo hydroxytiazolidín (Cys)). Tento intermediát sa zvykne vrátiť naspäť do pôvodného tvaru vypudením atakujúcej skupiny, keďže amid je preferovaný na základe voľnej energie (pravdepodobne kvôli silnej rezonančnej stabilizácii peptidovej skupiny). Ďalšie molekulárne interakcie však môžu spôsobiť, že amidová skupina nebude dostatočne stabilná - vtedy sa namiesto toho vypudí aminoskupina a vznikne ester (Ser/Thr) alebo tioester (Cys), ktorý nahradí pôvodnú peptidovú väzbu. Táto chemická reakcia sa nazýva N-O acylový posun.
Výsledná (tio)esterová väzba môže byť štiepená roznymi spôsobmi:
- Obyčajná hydroláza rozštiepi polypeptidový reťazec, kde sa štiepená aminoskupina stáva novým N-koncom. Tento typ reakcie prebieha pri vzniku glykozylasparaginázy.
- β-Eliminačná reakcia takisto štiepi reťazec, ale novým N-koncom sa stáva pyruvylová skupina. Táto skupina môže byť u niektorých enzýmov použitá na napojenie katalytického kofaktoru, hlavne u dekarboxyláz ako sú S-adenozylmetioníndekarboxyláza (SAMDC), ktorá využíva schopnosť pyruvylovej skupiny odťahovať elektróny.
- Intramolekulárna transesterifikácia, ktoré vedie k vetvenému polypeptidu. U inteínov je nová esterová väzba štiepená intramolekulárnym atakom asparagínu, z ktorého sa potom stane C-koncová aminokyselina.
- Intermolekulárna transesterifikácia môže viesť k presunu celého segmentu jedného polypeptidu na iný, ako je to u samospracovania Hedgehog proteínov.
Komprimácia sekvencie[upraviť | upraviť zdroj]
Komprimácie aminokyselinovej sekvencie, teda zmenšenie objemu sekvencie, je pomerne náročná úloha. Existujúce špecializované nástroje na komprimáciu sekvencie majú nízku úroveň kompresie v porovnaní s nástrojmi na komprimáciu DNA sekvencií, hlavne kvôli povahe dát. Aktuálne najlepší kompresor, ktorý komprimuje dáta bez straty informácie, je AC2.[8] AC2 využíva neurálnu sieť.
História[upraviť | upraviť zdroj]
Predstavu bielkovín ako lineárnych reťazcov α-aminokyselín zmienili takmer v ten istý moment dvaja vedci, na tej istej konferencii v roku 1902. Bolo to 74. stretnutie Spoločnosti nemeckých vedcov a doktorov, ktoré sa konalo v Karlových Varoch. Franz Hofmeister formuloval tento návrh ráno na základe poznatkov o biuretovej reakcii bielkovín. O niekoľko hodín neskôr ho nasledoval Emil Fischer, ktorý mal mnoho chemických detailov, ktoré podporovali model peptidovej väzby. Pred nimi tento návrh podal i francúzky chemik E. Grimaux, v roku 1882.[9]
Napriek dátam a neskôr i dôkazom, že proteolytické trávenie bielkovín produkovalo len oligopeptidy, predstava toho, že bielkoviny sú len lineárne nevetvené polyméry aminokyselín nebola okamžite prijatá. Niektorí rešpektovaní vedci, napríklad William Astbury, pochybovali o tom, že kovalentné väzby sú dostatočne silné na to, aby držali tak veľké molekuly pokope. Obávali sa toho, že náhodný pohyb molekúl spôsobený teplotou by postupne rozložil tvar týchto molekúl. Hermann Staudinger čelil podobným názorom v 20. rokoch 20. storočia keď tvrdil, že guma sa skladá z makromolekúl.[9]
Kvôli tomu vzniklo niekoľko alternatívnych hypotéz. Hypotéza koloidných bielkovín tvrdila, že bielkoviny sú koloidy zložené z menších molekúl. Táto hypotéza bola vyvrátená v 20. rokoch 20. storočia pomocou dvoch druhov meraní. Prvým z nich bola ultracentrifugácia, ktorú robil Theodor Svedberg, ktorý preukázal, že bielkoviny majú dobre definovaný a reprodukovateľnú molekulárnu hmotnosť: Druhými boli elektroforetické merania, ktoré robil Arne Tiselius, ktoré naznačovali, že bielkoviny sú zložené z jedinej molekuly. Druhá hypotéza bola cyklolová hypotéza rozvinutá Dorothy Wrinchovou, ktorá tvrdila, že lineárne polypeptidy podstupovali chemické cyklolové prešmyky C=O + HN C(OH)-N, ktoré viedli k previazaniu reťazca amidových skupín a tak tvorili dvojdimenzionálny materiál. Ďalší vedci predstavili iné hypotézy primárnych štruktúr bielkovín, napríklad diketopiperazínový model Emila Abderhaldena alebo pyrol/piperidínový model Troensegaarda v roku 1942. Aj keď týmto hypotézam nebola venovaná veľká pozornosť, tieto modely boli konečne vyvrátené, keď Frederick Sanger úspešne sekvenoval inzulín v roku 1955[10] a keď John Kendrew a Max Perutz určili štruktúru myoglobínu a hemoglobínu pomocou röntgenovej kryštalografie v rokoch 1957 a 1959.[11][12]

Primárna štruktúra iných molekúl[upraviť | upraviť zdroj]
Dá sa povedať, že každý heteropolymér s lineárnym reťazcov má primárnu štruktúru, analogicky k primárnej štruktúre bielkovín. Toto použitie je však vzácne v porovnaní s bielkovinami. U RNA, ktorá má širokú škálu sekundárnych štruktúr, sa väčšinou poradie nukleobáz označuje jednoducho ako „sekvencia“, podobne ako u DNA (ktorá zvyčajne tvorí dvojzávitnicu, ktorá má minimálnu sekundárnu štruktúru). Ostatné biologické polyméry, napríklad polysacharidy, majú takisto technicky primárnu štruktúru, ktorá je tvorená poradím podjednotiek, ale toto pomenovanie sa nepoužíva.
Vzťah k sekundárnej a terciárnej štruktúre[upraviť | upraviť zdroj]
Primárna štruktúra biologických polymérov do veľkej miery určuje trojdimenzionálny tvar (terciárnu štruktúru). Sekvencia aminokyselín v bielkovinách sa dá použiť na predpovedanie lokálnych vlastností, napríklad segmentov sekundárnej štruktúry, alebo transmembránových oblastí. Komplexita skladania bielkovín však aktuálne nepovoľuje určovanie terciárnej štruktúry bielkoviny len na základe jej primárnej štruktúry (poradia aminokyselín). Znalosť štruktúry podobných homológnych sekvencií (napríklad členov tej istej rodiny bielkovín) umožňuje vysokú presnosť predpovedí terciárnej štruktúry vďaka homológnemu modelovaniu. Ak je známa celá sekvencia bielkoviny, je možné odhadnúť jej biofyzikálne vlastnosti, napríklad izoelektrický bod. Rodiny bielkovín s podobnými sekvenciami sú určované pomocou klastrovania sekvencií a projekty štruktúrnej genomiky majú za cieľ vytvoriť set reprezentatívnych štruktúr sekvenčného priestoru možných neredundantných sekvencií.
Referencie[upraviť | upraviť zdroj]
- ↑ a b SANGER F. Advances in Protein Chemistry. Ed. M.L. Anson. [s.l.] : [s.n.], 1952. ISBN 9780120342075. DOI:10.1016/S0065-3233(08)60017-0 The arrangement of amino acids in proteins, s. 1–67.
- ↑ AASLAND, Rein; ABRAMS, Charles; AMPE, Christophe. Normalization of nomenclature for peptide motifs as ligands of modular protein domains. FEBS Letters, 2002-02-20, s. 141–144. ISSN 1873-3468. DOI: 10.1016/S0014-5793(01)03295-1.
- ↑ A One-Letter Notation for Amino Acid Sequences*. European Journal of Biochemistry, 1968-07-01, s. 151–153. ISSN 1432-1033. DOI: 10.1111/j.1432-1033.1968.tb00350.x. PMID 11911894.
- ↑ a b Hausman, Robert E.; Cooper, Geoffrey M.. The cell: a molecular approach. Washington, D.C : ASM Press, 2004. ISBN 978-0-87893-214-6. S. 51.
- ↑ a b c FASTA Substitution Codes [online]. web.cas.org, [cit. 2022-06-03]. Dostupné online.
- ↑ a b c d e f Codes Used in Sequence Description [online]. www.ddbj.nig.ac.jp, [cit. 2022-06-03]. Dostupné online. (po anglicky)
- ↑ a b c Codes and Abbreviations [online]. www.genome.jp, [cit. 2022-06-03]. Dostupné online.
- ↑ Silva M, Pratas D, Pinho AJ. AC2: An Efficient Protein Sequence Compression Tool Using Artificial Neural Networks and Cache-Hash Models. Entropy, April 2021, s. 530. DOI: 10.3390/e23050530.
- ↑ a b Fruton JS. Early theories of protein structure. Ann. N. Y. Acad. Sci., May 1979, s. xiv, 1–18. DOI: 10.1111/j.1749-6632.1979.tb14125.x. PMID 378063.
- ↑ WhatisBiotechnology • The sciences, places and people that have created biotechnology [online]. WhatisBiotechnology.org, [cit. 2022-06-02]. Dostupné online. (po anglicky)
- ↑ The Nobel Prize in Chemistry 1962 [online]. NobelPrize.org, [cit. 2022-06-02]. Dostupné online. (po anglicky)
- ↑ PERUTZ, M. F.; ROSSMANN, M. G.; CULLIS, Ann F.. Structure of Hæmoglobin: A Three-Dimensional Fourier Synthesis at 5.5-Å. Resolution, Obtained by X-Ray Analysis. Nature, 1960-02, roč. 185, čís. 4711, s. 416–422. Dostupné online [cit. 2022-06-02]. ISSN 1476-4687. DOI: 10.1038/185416a0. (po anglicky)
Pozri aj[upraviť | upraviť zdroj]
Zdroj[upraviť | upraviť zdroj]
Tento článok je čiastočný alebo úplný preklad článku Protein primary structure na anglickej Wikipédii.